
Icons of ID: Darwinian predictions and the Cambrian
Common, though persistent, misconceptions exist among ID proponents about the Cambrian Explosion, the Darwinian ‘tree of life’ and the appearance of phyla. I hope that my contribution will help put to rest some of these misconceptions.
These misconceptions include: 1) no Pre-Cambrian fossils 2) appearance of phyla are at odds with Darwinian predictions 3) the shape of the tree of life over time 4) no transitional fossils between phyla.
The confusion seems to have started with Art Battson, was popularized by Wells but continues with on Mark Hartwig and more recently with Salvador who posted the following picture (loosely based on the original work by Art Battson).

There are several significant problems with this picture. Lets start with the top left graph which shows the ‘Darwinian prediction’ for the origin of phyla. Looking at the bottom left figure, one notices that Darwin accepted extinctions and thus the suggestion that a ‘Darwinian prediction’ should be on of a continuously increasing number of phyla is wrong. The second problem is that the concept of phyla did not exist in Darwin’s time [1] and that Darwin was talking about the origin of species not necessarily higher taxa. Another problem is that Darwinian theory does not state anything about the speed with which phyla should arise.
Additionally, the top right picture which supposedly represents the actual data is misleading due to the compressed time scale and the discontinuities. The real question is: Is the origin of phyla at odds with Darwinian theory. As I will show, it isn’t.
The most problematic claim is that Darwinian theory predicts that phyla would arise as portrayed above. No evidence is provided as to how this prediction was made or derived. In fact I will show that the prediction is a strawman? By innappropriately comparing species trees (Darwin) with a Linnean hierarchy (phyla) a prediction is made that Darwin expected lower taxa to evolve into higher taxa. In otder to understand why this perspective is erroneous it is helpful to understand what the Linnaean hierarchy is all about/ While the Linnean concept of using nested hierarchies is compatible with an evolutionary perspective, a Linnaean categorization inevitably imposes equal distance to organisms in each category. Which means that a comparison between a Linnean hierarchy and an evolutionary tree can lead to misleading conclusions.
Linnaean Hierarchy
A good review paper on the Linnaean hierarchy and its importance versus other proposed phylogeny based hierarchies is Stems, nodes, crown clades, and rank-free lists: is Linnaeus dead? by Michael Benton in Biol. Rev. (2000), 75, pp. 633-648. A good essay which looks at the Linnean hierarchy and a recent proposal called Phylocode is ‘The future of biological taxonomy? Does the PhyloCode offer a viable alternative to traditional Linnaean taxonomy?’ by Jake Alexander.
As I have already shown the Linnaean hierarchy is artificial and indeed has been subject itself to many changes. While most biologists accept the concept of ‘species’ as real, higher taxa are less well defined.
Formally, there is limited advice about how to define taxa above the species level, such as genera, families, or orders, although in practice all systematists have used diagnoses consisting of characters.
That Linnaean nomenclature is articificial is shown by the various modifications made since Linnaeus first proposed his hierarchy.
Linnaean nomenclature has been modified both before the advent of cladistics (e.g. exclusion of polphyletic taxa, insertion of additional category terms, inclusion of fossil taxa), and after (e.g. exclusion of paraphyletic taxa, use of indented lists, sequencing, plesions). All these modifications have shared a common principle : that classifications should adhere ever closer to current knowledge of phylogeny, while at the same time remaining conservative.
Stems, nodes, crown clades, and rank-free lists: is Linnaeus dead? by Michael Benton in Biol. Rev. (2000), 75, pp. 633-648
In other words, these classifications are human constructs and no true classification exists:
Biological classifications, then, are necessarily entirely human constructs. There is no single, true classification inherent in Nature that is there to be discovered. In this paper, I use the term ‘ classification’ in two ways, as is common practice: first, a classification means an ordered list of species or higher taxa, and second, classification describes the process of achieving such an ordered list. The usages should be clear from context.
Stems, nodes, crown clades, and rank-free lists: is Linnaeus dead? by Michael Benton in Biol. Rev. (2000), 75, pp. 633-648
The Linnaean hierarchy is a hierarchy of taxonomic categories. It should not be confused with a hierarchy of taxa, the members of which may (or may not) be assigned to those categories. Therefore, even if taxonomic categories (kingdom: division/phylum: class: order: family: genus: species) were to be eliminated, hierarchies of nested taxa (Animalia: Chordata: Aves: Anseriformes: Anatidae: Branta: B. canadensis) presumably would not also be eliminated - at least not under an evolutionary world view. Regardless of the Linnaean hierarchy, the principle of common descent implies that the relationships among taxa will be nested and hierarchical.
The concept of a ‘phylum’ is an arbitrary division, or classification of a group of organisms. The moment you define a particular species under the Linaean hierarchy, you have to define its genus, family, order, class, phylum and kingdom. So contrary to some of the claims of ID, higher taxa arise first in a Linnaean hierarchy. But that’s because of the somewhat arbitrary and artificial nature of this hierarchy.
Because of these issues people over time has proposed (better) alternatives. In Proposal for a standardized temporal scheme of biological classification for extant species John Avise and Glenn Johns, propose ways to rectify the nomenclature of biological classifications.
The Linnaean system of classification (3) has served biologists for more than two centuries. Originally designed to catalogue diverse works of the Creator, the hierarchical categories in this ordering scheme later became interpretable as natural outcomes of the nested branching structures in evolutionary trees. Yet most classifications in current use continue to group species according to some unspecified mix of similarity by resemblance (phenetic grades) and similarity by descent (phyletic clades). Apart from this epistemological flaw, the kinds of empirical data used to recognize grades or clades vary greatly among organismal groups, with no explicit attempt to normalize assayed characters, to equilibrate taxonomic assignments, or even to adopt any universally standardized criteria for taxonomic ranking (4, 5).
Proposal for a standardized temporal scheme of biological classification for extant species by John Avise and Glenn Johns in Proc Natl Acad Sci U S A. 1999 June 22; 96 (13): 7358-7363
Based on the shortcomings of the Linnaean hierarchy, people have proposed PhyloCode. The advantage of PhyloCode is that it references phylogeny, the disadvantage is that this nomenclature is far less universal and stable than the Linnaean nomenclature. Not surprisingly the PhyloCode is not without its critics.
Linnaean nomenclature is stable enough to say what we know, flexible enough to accommodate what we learn; independent of specific theory, yet reflective of known empirical data; compatible with phylogenetic theory, but not a slave to it; particular enough for precise communication, general enough to reflect refuted hypotheses. LN is an effective international, inter-generational, and trans-theoretical system of classification that was forged and tested by those describing the earth’s biota, not touting political slogans. It has weathered more worthy adversaries than the Phylocode and will be in wide use long after the latter is a curious footnote to the history of taxonomy.”
and this one and hotly discussed on this TaxaCom archive
Shortcomings of Conventional Taxonomic Practice A primary limitation of conventional taxonomy is that extant taxa placed at the same Linnaean rank are not necessarily equivalent in age, diversity, disparity, or any other consistent property of their biology or evolutionary histories. Current taxonomic anachronisms communicate almost no information as to whether, for example, a rank such as genus, tribe, family, or order in mammals is equivalent to its counterpart rank in fishes, insects, or any other assemblage. Thus, the ranks in current use do little to aid, and indeed often may hinder, comparative evolutionary studies.
Proposal for a standardized temporal scheme of biological classification for extant species by John Avise and Glenn Johns in Proc Natl Acad Sci U S A. 1999 June 22; 96 (13): 7358-7363

(Click image to enlarge) Fig. 1 Examples of gross disparities of taxonomic assignments in current classifications. The phylogenies depicted, based on an integration of molecular and paleontological evidence, come from information in refs. 43 (a), 26 (b), and 44 (c). Proc Natl Acad Sci U S A. 1999 June 22; 96 (13): 7358-7363
The authors propose an alternative
Proposal for a Standardized Classification Scheme We propose that the approximate dates of nodes in evolutionary trees should be the universal criterion according to which taxonomic classifications above the level of biological species are erected. Decisions about the particular window of time to be associated with each taxonomic rank are arbitrary, but the conventions adopted should reflect some agreed-to consensus (7) among practicing systematists reaching this initial consensus may be the most difficult part of the entire endeavor).
Under this alternative the species of the Ciclids, primates and fruit flies would be placed in a hierarchical taxonomy as follows

(Click image to enlarge) Fig.3 The temporal-banding concept as applied to produce a time-standardized classification for the three groups of organisms in Fig. 1. Proc Natl Acad Sci U S A. 1999 June 22; 96 (13): 7358-7363
The shortcoming identified by these authors is also exemplified by the arguments by ID proponents as to the nature of the evolutionary tree. Once phyla arose, they are often portrayed as remaining ‘frozen’ in diversity, disparity or morphological distance.
Darwinian theory and the fossil evidence
As I said before: “While the Linnean concept of using nested hierarchies is compatible with an evolutionary perspective, a Linnaean categorization inevitably imposes equal distance to organisms in each category. Which means that a comparison between a Linnean hierarchy and an evolutionary tree can lead to misleading conclusions”.
A good example can be observed in the following graph:

Origin of phyla
Most recently, the DI Staff (DIS) (remember that they are arguing that origin of phyla etc is at odds with (Neo-)Darwinian theory, citing Valentine. Valentine is commonly cited by ID proponents and one may infer that the DIS agrees with Valentine’s interpretations:
DIS wrote:
More recently, in his 2004 book On the Origin of Phyla, paleontologist James Valentine evaluates various attempts to explain (or explain away) the origin of the body plans that arise in the Cambrian. He concludes that no current hypothesis provides a satisfactory account of the origin of the Cambrian phyla and that the problem of novel body plans remains unsolved–or, as he puts it “the underlying causes remain uncertain.”
versus
The title of this book, modeled on that of the greatest biological work ever written, is in homage to the greatest biologist who has ever lived. Darwin himself puzzled over but could not cover the ground that is reviewed here, simply because the relevant fossils, genes, and their molecules, end even the body plans of many of the phyla, were quite unknown in his day. Nevertheless, the evidence from these many additional souces of data simply confirm that Darwin was correct in his conclusions that all living things have descended from a commmon anscestor and can be placed within a tree of life, and that the principle process guiding their descent has been natural selection.
The data on which this book is based have accumulated over the nearly century and a half since Darwin published On the Origin of Species, some gradually, but much in a rush in the last several decades. I have been working on this book for well over a decade, and much of that time has been spent in trying to keep up with the flood of incredibly interesting findings reported from outcrops and laboratories. I am stopping now not because there is a lull in the pace of new discoveries (which if anything is still picking up), but because there never will be a natural stopping place anyway, and because the outlines of early metazoan history have gradually emerged from mysteries to testable hypotheses.
Valentine On the origin of phyla 2004, preface
The book by Valentine is a whopping 500+ pages but one would expect the DIS to at least have read the preface to the book.
Valentine hase been extensively quoted within ID’s Valentine on the Cambrian. One may wonder why Valentine’s more recent findings have not found their way onto this website.
PZ Myers discusses the ‘scholarship’ when DI describes the work by Valentine and others on the Cambrian explosion. In addition to relying on old papers, especially since in the last decade new fossils and phylogenetic data have added much to our knowledge of the Cambrian explosion, the DI seems to focus on ‘sound bites’ while missing the larger picture.
Darwin on his ‘tree of life’
The affinities of all the beings of the same class have sometimes been represented by a great tree. I believe this simile largely speaks the truth. The green and budding twigs may represent existing species; and those produced during each former year may represent the long succession of extinct species. At each period of growth all the growing twigs have tried to branch out on all sides, and to overtop and kill the surrounding twigs and branches, in the same manner as species and groups of species have tried to overmaster other species in the great battle for life. The limbs divided into great branches, and these into lesser and lesser branches, were themselves once, when the tree was small, budding twigs; and this connexion of the former and present buds by ramifying branches may well represent the classification of all extinct and living species in groups subordinate to groups. Of the many twigs which flourished when the tree was a mere bush, only two or three, now grown into great branches, yet survive and bear all the other branches; so with the species which lived during long-past geological periods, very few now have living and modified descendants. From the first growth of the tree, many a limb and branch has decayed and dropped off; and these lost branches of various sizes may represent those whole orders, families, and genera which have now no living representatives, and which are known to us only from having been found in a fossil state.

Click picture for larger version
I request the reader to turn to the diagram illustrating the action, as formerly explained, of these several principles; and he will see that the inevitable result is that the modified descendants proceeding from one progenitor become broken up into groups subordinate to groups. In the diagram each letter on the uppermost line may represent a genus including several species; and all the genera on this line form together one class, for all have descended from one ancient but unseen parent, and, consequently, have inherited something in common. But the three genera on the left hand have, on this same principle, much in common, and form a sub-family, distinct from that including the next two genera on the right hand, which diverged from a common parent at the fifth stage of descent. These five genera have also much, though less, in common; and they form a family distinct from that including the three genera still further to the right hand, which diverged at a still earlier period. And all these genera, descended from (A), form an order distinct from the genera descended from (I). So that we here have many species descended from a single progenitor grouped into genera; and the genera are included in, or subordinate to, sub-families, families, and orders, all united into one class. Thus, the grand fact in natural history of the subordination of group under group, which, from its familiarity, does not always sufficiently strike us, is in my judgement fully explained.
Chapter 13: On the origin of species
Darwin’s tree in other words is fully in line with the expectations and the data. To see this consider the bottom species marked capital A-L. While in Darwin’s graph they represent orders, they could equally well represent phyla. All these phyla by definition have to start at the bottom of the tree after the last common ancestor(s) until the species of those days had sufficiently diversified. In other words, that phyla arise at the bottom of the tree of life, is something easily overlooked. In this case, 12 phyla arose connected below with a common ancestor. So contraty to ID predictions, Darwinian theory is not accurately captured in the figure presented by Salvador.
Things do not get better though since the bottom right picture is again an oversimplification of the real tree of life. As Valentine shows, when taking into account the fossils and genetic knowledge of our times a picture emerges which is quite in line with Darwinian theory.
Linnaean Hierarchy
A Linnean hierarchy be definition has a species, genus, family, order, class, phylum and kingdom attached
- Levine, A., Review of Marc Ereshefsky, The Poverty of the Linnean Hierarchy, Quarterly Review of Biology, Spring, 2002.
- Brigandt, I. (2002) “The Linnean tradition under attack.” Review of ‘The Poverty of the Linnean Hierarchy. A Philosophical Study of Biological Taxonomy’ by Marc Ereshefsky. Metascience 11: 355-358.
If we had understood modern evolutionary synthesis during Linnaeus’ time, it is possible we would not have adopted the “binomial” convention; we would have been influenced by the knowledge that phylogenetic groupings occur at variable levels and no two are strictly comparable. Thus, the point where one draws the line between a generic distinction and a familial distinction is more or less arbitrary and is unlikely to be consistent between different lineages:
Common fallacies and misunderstandings
Common objections to the Cambrian explosion and the tree of life include the ‘top-down/bottom-up’ fallacy which is based on equivocation of two different kind of trees.
TOP-DOWN OR BOTTOM-UP
Some critics of evolution make much of the “top-down” versus the “bottom-up” pattern of appearance of higher taxa. That is, phylum-level diversity reaches its peak in the fossil record before class-level diversity, and the class-level diversity before that of orders, etc. These critics interpret this apparent “top-down” pattern as contrary to expectations from evolutionary theory. However, this pattern is generated by the way in which species are assigned to higher taxa. The classification system is hierarchical with species being grouped into ever larger and more inclusive categories. When this classification hierarchy is applied to a diversifying evolutionary tree, a “top down” pattern will automatically result. Consider species belonging to a single evolving line of descent given genus-level status. This genus is then grouped with other closely related lines of descent into a family. The common ancestors of these genera are by definition included within that family. Those ancestors must logically be older than any of the other species within the family. Thus the family level taxon would appear in the fossil record before most of the genera included within it. The “top down” pattern of taxa appearance is therefore entirely consistent with a branching tree of life.
Keith Miller on Wells
and
Continuing to move the shells, Wells invokes a semantic sleight of hand in resurrecting a “top-down” explanation for the diversity of the Cambrian faunas, implying that phyla appear first in the fossil record, before lower categories. However, his argument is an artifact of taxonomic practice, not real morphology. In traditional taxonomy, the recognition of a species implies a phylum. This is due to the rules of the taxonomy, which state that if you find a new organism, you have to assign it to all the necessary taxonomic ranks. Thus when a new organism is found, either it has to be placed into an existing phylum or a new one has to be erected for it. Cambrian organisms are either assigned to existing “phyla” or new ones are erected for them, thereby creating the effect of a “top-down” emergence of taxa.
Gishlick ICONS OF EVOLUTION?\ Why much of what Jonathan Wells writes about evolution is wrong
Dawkins:
Suppose you have a great oak tree with huge limbs at the base and smaller and smaller branches toward the outer layers where finally there are just lots and lots of little twigs. Obviously the little tiny twigs appeared most recently. The larger boughs appeared a long time ago and when they did appear, they were little twigs. What would you think if a gardener said, “Isn’t it funny that no major boughs have appeared on this tree in recent years, only small twigs?” You’d say he is stupid.
Richard DawkinsTHE “ALABAMA INSERT”: A STUDY IN IGNORANCE AND DISHONESTY
Richard Dawkins, THE “ALABAMA INSERT”: A STUDY IN IGNORANCE AND DISHONESTY Journal of the Alabama Academy of Science, Vol. 68, No.l, January, 1997. Franklin Lectures in Science & Humanities Auburn University April 1, 1996
Salvadore, I think you’re incorrectly assuming that what Darwin wrote is somehow at odds with what Dawkins wrote. Dawkins’ point is that phyla are named in retrospect, and they represent early branch points in evolution, some of which have left large groups of diverse survivors. More recent branch points, which have resulted in smaller amounts of divergence, naturally don’t get categorized as the most inclusive kind of group (a phylum) when they belong to older, more inclusive groups. So anyone who asks the question, “why are there no new phyla”, is failing to understand the basics behind phylogenetic classification. (The IDists didn’t actually come up with this misunderstanding, they borrowed it from Stephen J. Gould. Between him and the creationists, I don’t think IDists have ever come up with an original argument.)
All those pictures that you’ve posted have done is to take this misunderstanding and put it into visual form. There is no “Darwinian prediction” that new phyla should constantly arise; in fact, what you choose to call a “phylum” is arbitrary. We could call mammals a phylum if we wanted to, but then we’d have to call chordates a “super phylum” or some such. The charts also shows phyla arising straight out of the Cambrian without any change in morphology since. That’s just laughable. Few modern animals resemble any Cambrian animal other than by having a few diagnostic characters that define their phylum. (And again, those diagnostic characters are determined in retrospect.) Check out the worm-like Pikaia, the Cambrian’s representative chordate, and tell me how much it resembles an elephant or an ostrich:
Grape Ape at ARN

Although it has never been clear what hypotheses ID had in mind to explain the Cambrian explosion, it seems that when the veil of ignorance about the Cambrian Explosion is lifted, it appears to be well in line with the (neo-)Darwinian expectations. Has ID contributed in any manner to lifting this veil? On the contrary, only by representing the Cambrian Explosion as a strawman could ID even hope to make its case. And as is so common with arguments from ignorance, new data ended up showing ID to be wrong once again.
Salvador wrote:
Let’s try to be a little constructive about finding answers. I already admited it’s a guess, and I think a fair one. I’m offering to you to tell every one what you think the graph should look like if you find that guess so disagreeable. If you are saying the guess is inaccurate, suggest what would be a better model.
Or is it that you’ll have to admit, Darwinism never really gave predictions to begin with?
Interesting rhetoric, Salvador thinks the graph is a fair guess but seems unfamiliar with the writings of Darwin. IF he were correct and Darwinism never really gave predictions to begin with, then would the logical conclusion not be that the graph was ‘made up’? The real question, which the graph is trying to avoid by presenting a strawman, is : Is the data about phyla at odds with Darwinian theory. As I have shown it isn’t. The graph is a poor representation of factual data to promote a view of the Cambrian period which is at odds with the facts. And once again ID shows itself to be scientifically irrelevant, or in this case scientifically detrimental.
Salvador wrote:
So to be fair, are you aware of whether Darwin or people making extrapolations of his theory, made any prediction about the increasing or decreasing of Phyla?
The most parsimonious interpretation (to me) is to assume that Darwinism predicted 1 phylum to start off with, and then that phylum became many. The graph of “Darwinian Predictions” reasonably depicts that idea.
Has it occured to Salvador to read what Darwin had to say about this? Remember that the concept of phyla did not exist in those days and that Darwin mostly focused on species (On the origin of Species). Nevertheless it is clear that Darwin does include extinction thus any graph which fails to take into account extinction is at odds with Darwin’s work. I thought that Salvador was presenting a ‘prediction’, now it seems to be the ‘most parsimonious interpretation’…
Salvador wrote:
However, actual data does not yet agree with that view. It could be:
The transitionals existed, but we’ve just not found them
Some sort of Punctuated Evolution
Common descent is not how it happened
Something else
It’s likely 4.: The view is wrong. ID presents a strawman argument and even their portrayal of actual morphological distance is erroneous. That ID has chosen to present a ‘Darwinian prediction’ based on 1) assumptions that are not clearly specified and in fact appear to be erroneous 2) ignorance of what Darwin actually wrote, shows once again that ID is scientifically without much merrit.
No pre-Cambrian fossils
When Charles Darwin wrote On the Origin of Species, he and most paleontologists believed that the oldest animal fossils were the trilobites and brachiopods of the Cambrian Period, now known to be about 540 million years old. Many paleontologists believed that simpler forms of life must have existed before this but that they left no fossils. A few believed that the Cambrian fossils represented the moment of God’s creation of animals, or the first deposits laid down by the biblical Flood. Darwin wrote, “the difficulty of assigning any good reason for the absence of vast piles of strata rich in fossils beneath the Cambrian system is very great,” yet he expressed hope that such fossils would be found, noting that “only a small portion of the world is known with accuracy.”
Since Darwin’s time, the fossil history of life on Earth has been pushed back to 3.5 billion years before the present. Most of these fossils are microscopic bacteria and algae. However, in the latest Proterozoic - a time period now called the Vendian, or the Ediacaran, and lasting from about 650 to 540 million years ago - macroscopic fossils of soft-bodied organisms can be found in a few localities around the world, confirming Darwin’s expectations [uref]Introduction to the Vendian Period [/ref]
To learn more about Vendian fossils see Learning About the Vendian Animals or the exhibit at (the Paleontology Museum in Oslo (in Norwegian).
Disparity precedes divergence
Gould discussion
Gould pillories Walcott for his pragmatic inclusion of the Burgess Shale taxa into broadly defined phyla - ‘shoehorning’ in Gould’s terminology. But, here, Gould (also Mayr 2001, see p. 209) has forgotten a principle which is so fundamental it has become hackneyed: man classifies; nature does not. The only taxonomic rank possessing any degree of objective ‘reality’ is the species. To approximate disparity by counting phyla is to embark upon an extremely suspect enterprise.
and
Even if we confine ourselves to considering Phanerozoic animals, Gould is wrong. If we play Gould at his own game and approximate disparity by counting orders and higher taxonomic groupings (order and greater rank) within phyla, we find that:
* Brachiopoda achieve their maximum ‘disparity’ in the Devonian and Carboniferous periods; * Mollusca in the Cretaceous; * Echinodermata in the Ordovician; * Hemichordata (if this is in fact a monophyletic group) in the Ordovician; and * Chordata in the Carboniferous and Permian.
(Data after Rich et al. 1996.)
Finally, the pervasive pattern of geologic succession is systematically backwards from that predicted by the theory. Darwinian theory predicts that the gradual accumulation of minor evolutionary change and the increasing diversity of the lower taxa should ultimately produce the profound differences among the major body plans and the disparity of the higher taxa. Diversity should precede disparity. Geologic succession reveals the opposite: disparity precedes diversity. The major themes or body plans appear suddenly in the history of life only to be followed by variations on these pre-existing themes. The natural history of life on earth is systematically top to bottom, not bottom to top as Darwinian theory predicts.
Art Battson On the Origin of Stasis, Access Research Network Library File
The origin and differentiation of major clades is often assumed to have occurred in tandem with the ‘explosion’ of fossil evidence of diverse morphologies (‘disparity’) at the base of the Cambrian. Evidence is presented that this was not the case. Biogeographical and morphological differentiation among the earliest trilobites reveals incompleteness in the known early Cambrian record; similar evidence can be accrued for other major groups. Phylogenetic analysis proves the likelihood of ‘ghost’ lineages extending into the Precambrian. The important events in the generation of clades were earlier than the Cambrian ‘explosion’, at which time the groups become manifest in the fossil record. It is likely that the important phylogenetic changes happened in animals of small size; sister taxa of major groups are shown to be small animals. Decoupling cladogenesis from the Cambrian ‘explosion’ removes the necessity of invoking unknown evolutionary mechanisms at the base of the Phanerozoic. Genes controlling development may have played a role in generating new morphologies, through heterochrony for example, in the early differentiation of metazoan body plans.
The Cambrian evolutionary ‘explosion’: decoupling cladogenesis from morphological disparity by R.A. Fortey, D.E.G. Briggs and M.A. Wills in Biological Journal of the Linnean Society (1996), 57: 13-33.
- The basic hierarchy as formulated by Linneus, is as follows: Imperium (“Empire”), Regnum (“Kingdom”) Classis (“Class”), Ordo (“Order”), Genus, Species, Varietas (“Variety”) . Over time ‘Imperium’ was dropped and variety lost much of its relevance. Two new ranks were added: phylum (between kingdom and class) and Family (between order and genus). The modern Linnaean System is based on a simple hierarchical structure in which organisms are sorted using the nomenclature of Kingdom, Phylum, Class, Order, Family, Genus and Species.
Other websites that use these graphics or similar arguments
Fossils Reject the “Tree of Life” at DarwinismRefuted.com
Casey Luskin et al Icons Haven’t Lost Their Touch
Misleading: Regardless of what appears later in the fossil record, the bottom line is that almost all phyla appear in the Cambrian without any previous animal fossils to account for their supposed evolutionary origin.
The Creation Debate: The Fossil Record – Part One by Dr. John Ankerberg, Dr. Duane Gish, Dr. Kurt Wise
I believe that I may have located the orign of at last one of the graphs. It is from On the Origin of Stasis (Part I) By Art Battson found on ARN.
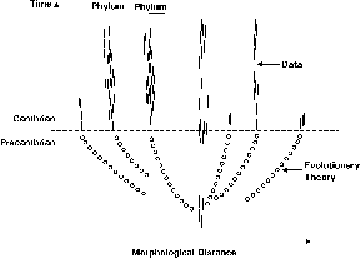
Darwinian Theory vs. the Fossil Record
Darwinian Theory attempts to explain the common ancestry of all species through the gradual transformation of major body plans. This theory is in opposition to the fossil evidence and the pervasive patterns of natural history. An estimated 75 to 100 phyla appear explosively at the base of the Cambrian. Fossil evidence suggesting their common ancestry is not found in Precambrian rocks. A General Theory of Macrostasis is needed to explain the fossil data and the stability of the higher taxa.
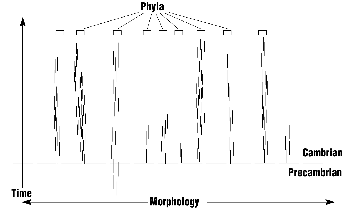
The Fossil Evidence
Contrary to both Darwinian gradualism and punctuated equilibria theory, the vast majority of phyla appear abruptly with low species diversity. The disparity of the higher taxa precedes the diversity of the lower taxa.